 ≫
古代で遊ぼ ≫
書庫(参考図と仮説) ≫
DNA分析による日本人形成 工事中
≫
古代で遊ぼ ≫
書庫(参考図と仮説) ≫
DNA分析による日本人形成 工事中
DNA分析では日本人の誕生をどう考えることができるのでしょうか。
「DNA人類進化学/宝来聴著/岩波書店」を参考にした仮説をたててみます。
DNAと日本人1 DNA分析とは、その現状
DNAと日本人2 宝来聴氏のDNA分析による日本人
DNAと日本人3 ユーラシアンとオーシャノイド仮説
DNAと日本人4 日本の先住者と縄文への仮説
DNAと日本人5 縄文と日本人を構成するDNA
DNAと日本人6 アジア人に欠失するDNA
DNAと日本人7 JCウイルス分布と日本人
DNAと日本人8 総括
DNAと日本人番外編(血液型)
(薄墨の背景になっている部分はなんらかの根拠のある情報を示します、その他は仮説)
DNAと日本人1
人間のDNAには約30億個の「塩基」(遺伝情報)があるそうですがまだ未解読です(核DNA)。
しかし、人間にはもうひとつミトコンドリアDNAというのがあります。
16569個の「塩基」と37個の遺伝子を持つDNAで、可読済みです。
その塩基の配列は長い時間の間に少しづつ変化しながら子に伝わっていてこれを塩基置換といいます。
塩基置換(≒突然変異)が一定の確立で発生する、と仮定することで時間経過を示す指標になるのだそうです。
(ただし、一定の確立で起きるとは限らない可能性も指摘されるようです)
ミトコンドリアDNAには大きな特徴がふたつあります。
ひとつは塩基置換の速度が核DNAより10倍近く早いこと。
例えばチンパンジーと人間の核DNAの差(違い)は1%なのですが、ミトコンドリアDNAでは9%の差になるのだそうです。
物差しの目盛りを細かくできるということでしょう。
もうひとつは母系にしか伝わらないDNAだということです。
男性にもミトコンドリアDNAはありますが、合体したとき男性のDNAは消滅するのだそうです。
核DNAの場合は母と父の複合体になるために、数世代を経ただけでその組み合わせはねずみ算で増え、10世代遡るだけで祖先は2の10乗=1024人も存在することになって分析不可能となってしまう。
しかし、母系だけにしか伝わらないミトコンドリアDNAなら1本道で分析ができます。
しかし、これは重大な欠陥を含むことになります、母方の系統しかわからない。
日本人、韓国人、中国人の293人のサンプルのDNA分析では下図のような関係があるそうです。
数字は関連の深さを表します。破線はほとんど関連が認められないことを示します。
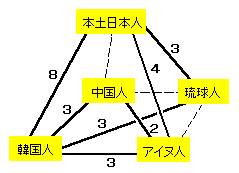
日本文化のほとんどが中国文化の影響下にあるのに、血縁的に中国人と無縁・・ほんとかな??
縄文人の母と中国人の父の間に子ができたとき、その子のDNAを分析しても中国人の父は発見できない。
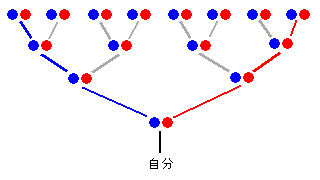
4世代前の祖先は16人います。
DNA分析でわかるのは右側の母系(赤線上)の女性だけで、他の祖先15人は発見できない。
それではどうにもならないようですが、それはある個人のDNAだけを遡った場合です。
サンプルをたくさん集めれば、右側ではない女性につながる人もいるわけで、統計処理すればそのサンプル集団全体としてある方向性が推測できるわけです。
個人分析では垂直方向の情報が得られ、集団では水平方向の情報が得られる、そんなイメージでしょう。
(この部分はちょっとむずかしい概念、表現が正しいかどうか自信なし(^^;)
例えば戦争で征服された国には征服者を父にする子が少なからず生まれるでしょう。
その子等が後に有力氏族になったとしても、その父方の祖先である人々の存在を検出できない。
文化や容貌に明確な変化があったにもかかわらず、DNA分析ではそれがわからない。
現状のDNA分析は民族移動のごとき大規模な女性を含む混合があった場合に、それを検出できる。
そのように理解しています。
最近、男性にしか伝わらないY染色体のDNA分析法が登場し始めているようですが、まだ不安定要素があるようです。
これの完成を期待するところです。
DNA分析の理解についてはこちらの質疑応答が有益と思います。ご参照ください。
以下は自分の理解が間違っている可能性もあります、その場合はご容赦。
DNAと日本人2 (topへ戻る)
DNAのある種のアミノ酸は内部時計を持っていてそれを分析すると分岐(突然変異)のあった年代を求めることができます。
下図はDNA人類進化学に載っているそのその簡略図です。
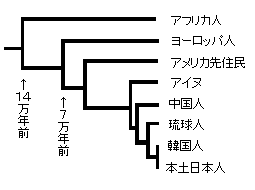
(年代は遺伝距離という数値で書かれていますが、実年代に換算してあります)
14万年前頃にアフリカから出発した人々がいた。
それがさらに7万年前に分岐して一方はヨーロッパ人になり、一方はモンゴロイドとなった。
DNAを遡ってゆくとみなアフリカ人につがってゆくわけで、人類アフリカ単一発生を示すわけです。
日本人の起源を考える場合に古モンゴロイドとか原モンゴロイドが日本にまずあって、といった論調をみかけますが、これは人類が各地で誕生進化したとする説に基づくものでしょう。
この説はDNA分析からは否定されます、とりあえずは。
日本人の基本DNAには異なる2系列が混在していて、その分岐は12万年前に遡るらしい。
12万年前ということはほとんど最古に近い人々を祖先にしているということになります。
これは単純分岐だけでは説明できないのだそうです。
順次分岐してきて日本人が登場したのではない、ということです。
下図は静岡県三島の116人のDNAを分析した結果で、62タイプがあったそうです。
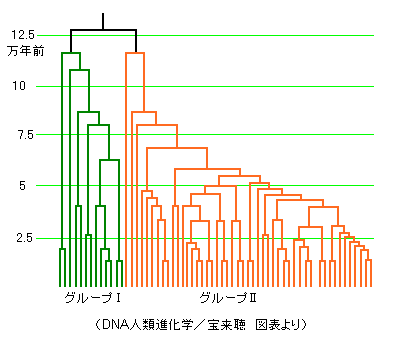
12万年前の分岐以降を緑と橙に色分けしてあります。
日本人には12万年前に分岐した2系列の人々が混在している、ということ。
宝来聴氏はグループⅠ(緑)とグループⅡ(橙)と称しています。
モンゴロイドが7万年前に登場したとすると、三島の日本人はそれより5万年も前の人間を祖先にしているということになります。
日本人以外のアジア人もグループⅠとⅡにわかれることがわかっているそうです。
すなわちアジア人はモンゴロイドが登場する以前の人々を祖先にもつ、ということです。
下図は埼玉県浦和で発見された縄文人骨(BC4000)とアジア人やヨーロッパ人との比較データです。
この縄文人は先の三島でのグループ分けにあてはめるとグループⅡに属しているのだそうです。
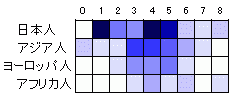
数字はDNAが縄文人と異なる部分がなければ0、1カ所異なるなら1、2カ所で2、数字が大きくなるほど異なる部分が増えた状態を示し、その状態での比較になります。
色を塗った部分はそれぞれの状態での縄文人と一致する人数で、色の濃いほど一致人数が多いことを示します。
浦和の縄文人と完全一致したのはインドネシア人とマレー人で2人いたそうです(0とアジア人の交点です)。
むろん縄文人全般がインドネシア人やマレー人と同じだということではなく、浦和の縄文人(個人)の場合はそうなるということです。
完全一致ではないけれど準じての一致では圧倒的に現代日本人に類似です。
次いでアジア人の類似でこれは当然と思えますが、アフリカ人よりヨーロッパ人と近似が多いことに注目です。
また、現代日本人でヨーロッパ人に近い人々は、青森28%、静岡18%、沖縄5%だったそうです。
列島の北部にヨーロッパ人と近い人が多いということです。
日本周辺の人々の同様分析がほしいですが、あいにくデータがありません。
北海道のBC1000頃の縄文人3体と近世アイヌの骨6体の分析も行われています。
結果はその縄文人は近世アイヌの一部と近似であり、同時に現代人の一部と東南アジアの一部とも近似という結果だそうです。
ま、これだけでは日本人みな兄弟ということしかわかりませんけど(^^;
アイヌとヨーロッパ人の近似度はどんなものなのかなあ、これもデータがありません。
次にアフリカ人、日本人、ヨーロッパ人に大別した場合の分岐系統樹をご覧ください。
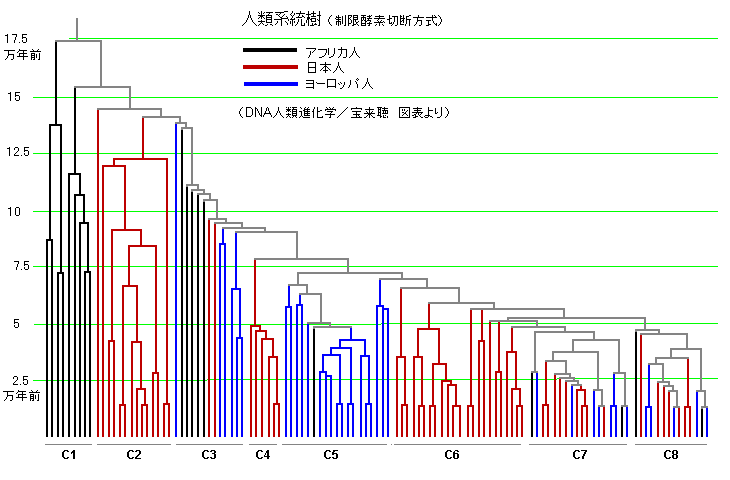
赤が日本人から検出された系統、青が同じくヨーロッパ人、黒がアフリカ人から検出された系統を示します。
日本人に多いのがC6グループで5万年前あたりでの分岐、次いで多いのがC2グループで15万年前あたりの最古の分岐の人々。
ヨーロッパ人は7万年前あたりに分岐したC5グループがほとんどを占めています。
分岐年代からみると三島の分岐図のグループⅠが本図のC2、グループⅡがC4、C6などに相応すると解釈しています。
日本人には現在のヨーロッパ人が登場する以前の人々を祖にする人々が多数含まれているということです。
DNAと日本人3 (topへ戻る)
単純分岐の考え方だけではここまでのいろいろを説明できないようです。
そこで、混血の概念を拡大した仮説を立ててみます。
人間の移動(行動)は環境に支配される。
少なくとも縄文時代あたりまでの人々の移動の引き金はすべて気候だと考えています。
その一番は食糧、食べ慣れた食糧を得るために住み慣れた環境と同じ地域へ移動してゆく。
むろん人間以外の動植物も同じです。というよりその動植物を追って人間が移動する。
寒冷化では南下する。
温暖化では北上する、山岳へ移動することもあるでしょう。
アフリカをでた人類が12万年前あたりで一方は内陸を東進しもう一方は沿岸部を東進していた。
内陸を東進した人々をユーラシアンと称しておきます。
その後、徐々に進行する寒冷化で寒冷適応の狩猟文化を発達させた。
強い日射から肌を守る必要が薄れてメラニン色素が減り肌は白くなる。
厳しい環境ゆえに道具(石器)の発達がうながされるでしょう。
沿岸を東進した人々をオーシャノイドと称しておきます。
赤道に近く最寒冷時代でも温暖な地域です。
温暖適応文化(農耕)を持つようになり肌は黒いままでしょう。
東南アジア到達時には赤道付近に南下し海に囲まれて海洋文化も持つようになった。
下図は南極の氷と福井県三方湖の花粉分析による気候変動推定図です。
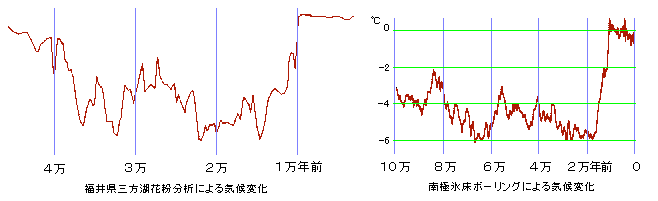
南極の気候が各地の気候に一致するわけではありませんが、同じ傾向を示すでしょう。
10万年~9万年前にウルム氷河期の前兆となる寒冷化がはじまり、寒暖をくりかえしながら7万年前からウルム氷河期の最初の寒冷ピークに突入しています。
福井県の花粉分析では4.3万年~3.8万年前に間氷期(若干の温暖、多雪湿潤)がはさまるようで、これはギリシャでの分析でも同様だそうです。
そして2万年前に再び寒冷ピークとなり、以降は急激に温暖化してゆきます。
仮説1
7万年前頃からのウルム氷河期本格化でユーラシアンは南下し、千年単位で繰り返される寒暖が人々を南北に揺り動かしながら寒冷かつ内陸に適応したユーラシアンと温暖適応あるいは海洋適応のオーシャノイドが混血していった。
モンゴロイドは単純分岐で登場したのではなく、別系に分かれて数万年を経た人々の再混血で登場した。
南北や東西で様相の違う混血になると思います。まずは西アジアからになるでしょう。
5万~2万年前の推定図参照
(スンダ:マレーシア、インドネシア、ボルネオ島が陸続きになっていた大陸)
(サフール:オーストラリアとニューギニアが陸続きになっていた大陸)
ヒマラヤ山脈は巨大な壁です。
南下と北上による混血がここでは阻止されて、南のインド亜大陸ではオーシャノイドがそのまま残り、北のタクラマカン砂漠(草原)付近ではユーラシアンがそのまま残る可能性が生じそうです。
東アジアでは長江と黄河が東西に移動を促進させ、メコンは南北の移動を促進させた。
そしてメコンを越えてスンダの東岸に到達した人々は海を渡る技術を持つようになっていった。
そして1.5万年前に至って世界は急激な温暖世界へ突入する。
数万年の寒冷時代に順応していた人々、平均気温で8度も気温が上昇したら動植物もこぞって涼しい場所を探すと思います。
かっての温暖地帯である東南アジアは熱帯雨林化し、それまでの文化は衰退していった。
太陽がたくさんあったとか、太陽の数を減らして涼しくなったという神話が世界中にあります。
この温暖化によって涼しさを求めた時の印象、これが生み出した神話ではないかと思っています。
オーシャノイドは北のユーラシアンの地域へ、ユーラシアンはより北へ北上していった。
あるいは山岳地へも移動していったでしょう。
各地でそれまで以上の急激な混血が生じ、新しいモンゴロイドの登場が加速された。
混合は刺激ともなって新しい文化の発達も促されたのではないかと思います。
北緯30度付近、チグリスユーフラテス、インダス、ガンジス、長江や黄河、その周辺で。
狩猟と農耕の合体=動物を作る=遊牧も登場した。
DNAと日本人4 (topへ戻る)
北アメリカで発見されたケネウイックマン(スピリットケープマン)はコーカソイドに類似の頭骨の持ち主です。
北上してきたモンゴロイド勢力と温暖化に追われて、陸続きとなっていたベーリング海峡を渡ったユーラシアンであると考えることができます(BC10000頃か)。
この発見でアメリカ先住民とはなにかの問題で議論が生じているようです。
そしてさらなる温暖化で勢力を増したモンゴロイドがアメリカ大陸へ渡ってゆく。
こちらは古来からの説におけるアメリカ先住民とされる人々の祖先。
西アジアや中近東の人々は12万年前あたりの、ユーラシアンでもオーシャノイドでもない人々の子孫。
そこに南北からの人々が再び混じったのが後の中近東イラン系の祖先ではないかと考えています。
(中近東の人々のDNA分析ぜひ期待したいです)
東ヨーロッパやシベリアまでオーシャノイドは北上せず、ここにはユーラシアン(の資質)がそのまま残った。
それがヨーロッパ人などいわゆる白人系、コーカソイドの祖先。
現在のヨーロッパ人はDNAの多様性が少ない人種だそうです。
二人だけから人口が増えたのならその二人のもつ多様性しか子孫は持たないわけで、容姿、文化や言語に共通性が多くなります。
ヨーロッパ人に多様性が少ないのは、ある時期に絶滅寸前となり、そこから再び人口が増えたため、という論があります。
ヨーロッパのネアンデルタール人はウルム氷河期が最寒冷となる3万年前頃に滅びたそうですが、同様に寒冷によって絶滅寸前から生き延びて復活した人々の子孫が現在のヨーロッパ人なのかもしれません。
2章の人類系統図のC5グループにヨーロッパ人が集中しているのはそれを示すのかもしれません。
なお、ネアンデルタール系が単に滅びたのではない可能性もあります。
中近東の4万~3万年前のアムッド人、スフール人、タブーン人などはネアンデルタール系に分類されるようですがホモサピエンスとネアンデルタールの中間的な特徴をもっています。
南下したネアンデルタール系の人々とオーシャノイド系の人々が混合してゆけばそういうことも生じるはずです。
イタリアで発見されたウルム氷河期のグリマルディ人では上層の骨はフランスのクロマニョン人と類似だが下層の骨ではニグロイドの特徴があるそうです。
またフランスのシャンスラード人ではクロマニヨンに類似だがエスキモーに見られる特徴を持つそうです。
これらの状況と仮説を加味して宝来氏のDNA分岐図を書き替えたものが下図です。
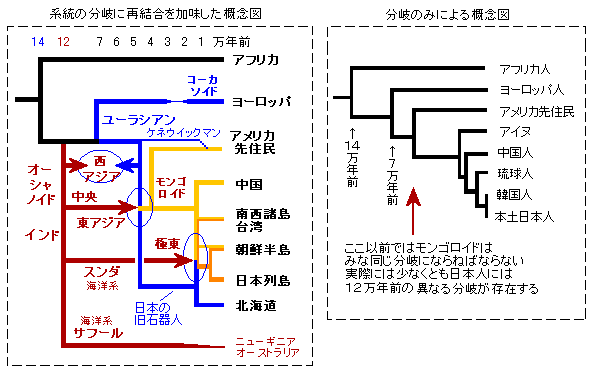
青丸が主要な混血ですが、その年代はいいかげんです。
仮説2
BC20000頃の日本にいた旧石器人はユーラシアン。
骨の発見を期待するところです。
沖縄の湊川人はBC16000で微妙な年代です。DNA分析期待したいです。
ユーラシアンが南西諸島に渡る必然性はないとみて、オーシャノイド系譜であろうと予測しておきます。
野尻湖でナウマン象を狩った人々の使った旧石器は北海道系です。
瀬戸内から出土する旧石器は沿海州系の石器です。
瀬戸内でナウマン象を狩った人々は朝鮮半島経由の南回りでやってきた人々なのでしょう。
日本には2系列の旧石器が出土しており、その接点が中部山岳地帯。
同じユーラシアンでも西日本と東日本では違いがあったかもしれません。
そして温暖化がはじまり海面上昇でスンダ大陸とサフール大陸が海没してゆきます。
人々はそこから脱出せねばなりませんが、5000年でおおよそ100mほどの海面上昇とみれば1年あたり2cm。
親の代の土地が子の代で少し減る、数世代後には移動しなければならないといった速度で、パニックになるわけではなく移動先をじっくり考えることはできたでしょう。
仮説3
オーシャノイド、海洋系の人々は東シナ海沿岸あるいは南西諸島伝いに日本へやってきた。
鹿児島(BC10000頃)や小笠原(年代不明)から丸木船をくりぬくのに最適の形状の丸ノミ型の磨製石斧が出土しています。
古黄河と古長江の冷水は古九州沿岸と古南西諸島周辺に流れ込み、古黒潮と混合しています。
この海域は大漁場になり、海洋民も集まってくる。
沿岸に拠点ができ、陸系の人々との接触も増えるでしょう。
中国大陸内部ではあっちこっちから全部が混じるのでわけがわからない状態です(^^;
BC8000頃には日本と大陸は海で切り離されますから、それ以降での列島では海洋民との接触がほとんどになるでしょう。
朝鮮半島の南岸もほぼ同様です。
グループⅠの分岐は古い、こちらはユーラシアンの系譜のDNAの持ち主、日本の先住旧石器人でシンプルなDNAの持ち主。
グループⅡはオーシャノイド系譜のDNAの持ち主、東南アジア、中国での分岐を含んで多様なDNAの持ち主。
温暖化でグループⅡの人々は北海道付近まで北上し、グループⅠと混合していった。
仮説4
古いユーラシアン系譜とオーシャノイド系譜が新しい時代(BC10000頃)に混血して登場した最新版のモンゴロイド、それが縄文人。
古くて新しい、だから日本人には12万年前に分岐したグループⅠとグループⅡの2系列のDNAが混在する。
DNAと日本人5 (topへ戻る)
中国人66人、琉球人50人、韓国人64人、本土日本人62人、アイヌ51人のDNA分析結果の概要がDNA人類進化学にあります。
中国人では66人中の固有タイプは54種、共通タイプは4種。
琉球人では50人中 〃 40種、 〃 4種。
韓国人では64人中 〃 36種 〃 13種。
本土日本人では62人中 〃 41種 〃 12種。
アイヌ人では 51人中 〃 18種 〃 7種。
詳細データはありませんが日本人登場への重要なヒントです。
中国人では8割が異なるDNAタイプで多彩、複雑な混血が多かったことを示すものです。
アイヌ人では異なるDNAタイプが3割しかありません。
他のタイプが消滅したためかもしれませんが、もともと混血が少なかったと考えられます。
本土日本人と韓国人はその中間的な様相を示しています。
まず日本全土にユーラシアンありき。
温暖化による南方系の人々の渡来と北上で本土は縄文化してゆきます。
東アジアのモンゴロイド参照)
大陸系モンゴロイドが温暖化で勢力を拡大してアメリカ大陸へ渡ったとき(BC8000頃)、北海道はすでに海で切り離されています。大陸系モンゴロイドは北海道にはやってこなかった。
縄文が北上し、北海道に達した頃に温暖化が終わります(BC3000頃)。
北海道の縄文化はそこで停止し混血も停止した。
北海道は微妙なバランスポイント(空白地帯)になっていたわけで、ここに最古のユーラシアンの資質(グループⅠ)が濃く残った。
これがアイヌの人々の祖先だと考えています。
北海道の状況推定図
沖縄での分析ではグループⅠに含まれる人々はわずかに5%だったそうです。
沖縄にユーラシアンが達するには相当な距離の海を渡らねばなりません。
内陸系の人々がここにやってくることはなかった、それを示すものと思います。
沖縄のBC16000頃の湊川人はオーシャノイド系譜の海洋系の人々。
湊川人は男子で身長153cmほど、骨格から上半身は華奢で力仕事には向いていないが下半身はがっちりしているのだそうです。
縄文人一般に同様の小柄ですから、縄文人はこの流れの影響を強く受けた可能性がみえます。
ただし、湊川人は縄文人ではありません、縄文の祖先の一方となった人々です。
沖縄は中国に次いで異なるDNA系譜をもつ人々が多い。
これは縄文中期~末期に長江付近の文化が発達し、その系譜の人々と文化の流入が継続し、弥生から近世に至る間でも中国や東南アジアとの交流が深いはずで、この影響を受けているものと考えられます。
したがって湊川人と現在の沖縄の人々とのDNAはそれほど近くはない、そう予測しておきます。
(骨からDNA分析ができるようになることを待ちます)
日本全土に内陸系(北方系)のユーラシアンがまずありき、これが日本人の第1基層。
DNAでのグループⅠです。
これにBC12000頃からオーシャノイドの文化と人々が入って縄文が誕生、これが日本人の第2基層。
DNAでのグループⅡです。
日本海に暖流が流れ込んで日本は多雪気候となり落葉広葉樹が大発達し、そこから得られる豊富な食料によって東日本の縄文人口は激増する。
縄文は大陸系のモンゴロイドと資質は類似になっても異なる文化をもつことになります。
最温暖期のBC4000頃には、沖縄や九州などの温暖な沿岸地域に東南アジアや東シナ海沿岸の最新文化(照葉樹林文化と漁労文化)を持つ人々が渡来し、各地の沿岸部へ展開していった。
多彩なDNA構成をもつ人々です(これもグループⅡ)。
話が前後しますが、BC15000以降の温暖化によって文化と人の栄える位置=最適環境の位置は赤道付近から北緯30度前後の大河川流域にシフトしていった。
東アジアでは長江流域。
対して東南アジアは熱帯化によってかってのオーシャノイド文化は衰退してゆきます。
BC2000頃以降では発達した各地の文化(文明)の交流や抗争によって男性だけによる混血も増えると思います。
中央アジアの寒冷型の大森林は大草原へ変身して遊牧文化が登場し、馬を用いて東西の移動が高速かつ容易になります。
こうなると母系しかつかめないDNA分析では状況把握は無理でしょう。
弥生時代の大陸系渡来者の西日本での拡大、農耕による人口の激増、これが第3基層ということになりますが、この時代では男性だけによる混血が増えるはずで、現状のDNA分析による考察は無意味だと思います。
男性だけに伝わるY染色体のDNA分析の進展が待たれるところです。
DNAと日本人6 (topへ戻る)
アジア人にはDNAの一部(9塩基)が欠失している特徴があるそうです。
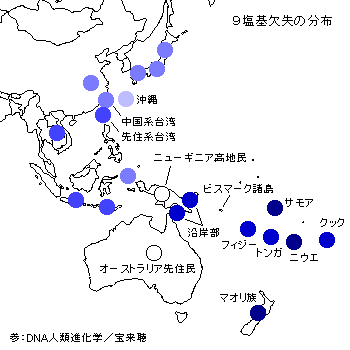
欠失を持つ人の多い地域が濃い青です。
ポリネシアとニュージーランドでは100%近くの人々がこの欠失をもつそうで、ラピタ文化の範囲と重なってきます。
(BC2000~BC1500頃にニューギニアの東のビスマーク諸島に突然登場した文化、周辺にはない土器文化をもつ)
日本、中国、東南アジアの人々の場合はおおよそ30%ほどがこの特徴を持ち、オーストラリアとニューギニア高地にはこの欠失がありません。
欠失を持つのはグループⅠだけなのだそうで、北方内陸系しか持っていない形質(ユーラシアン)がなぜポリネシアに登場したのでしょうか。
持論でゆけばユーラシアン系譜とオーシャノイド系譜の混合からこの欠失を濃厚に持つ人々が周囲に拡散したと考えることができます。
すなわちその発祥はユーラシアンが存在した地域ということになり、大陸沿岸が濃厚となります。
ポリネシアなどの人々の登場は新しい時代のようで、ラピタ文化の人々がその人々であるとみるなら、拡散年代はBC2000以降となります。
広大な範囲に拡散していますから海洋も得意にする有力な文化を持つ人々だったはずです。
長江良渚文化はBC2000頃に長江の大洪水で滅亡します。
ここから周辺に散った人々がその最有力であると考えています。
中国史書の尭舜禹時代に登場する三苗、それがこの欠失を持つ人々だったのではなかろうか。
後の黄河系文化からは悪役扱いをされていますが、BC5000~BC2000頃の長江文化を担った人々です。
欠失をまったく持たないニューギニア高地やオーストラリアの人々はその人々と接触しなかったのでしょう。
ニューギニア以東(ポリネシア)では先住人口が少なく、新たな渡来者もいなかったために欠失をもつ形質が100%近くで残ったのではないでしょうか。
ポリネシアの人々の容姿はミクロネシアやメラネシアとは異なってモンゴロイドに近い。
ポリネシアの人々は大柄ですが、中国南部には三苗伝承と同時代に防風氏という巨人族伝承もあります。
その人々が九州や朝鮮半島南岸に渡来し、BC2000以降の寒冷化で衰退していた縄文文化に新文化をもたらし、
プレ弥生文化の担い手(海洋系文化→記紀にいう海神など)になったと考えています。
タイ北部のノンノクタ遺跡やバンチェン遺跡などではBC2000~1000頃から青銅生産が始まります。
中国での青銅器登場より500年ほど遅いだけ、殷の青銅器と同時代です。
これも長江文明滅亡から周辺に散った人々によって開始されたと考えることができます。
タイ東北部のバンドンブロン遺跡では鉄生産がBC300~BC200頃に始まっています。
鉄生産でも中国での鉄生産と数百年の差しかありません。
これらの文化の流路、9塩基欠失の人々はBC2000以降のアジア情勢の重要なキーワードになると考えています。
なお、バンドンブロンの製鉄は粒状水酸化鉄を炭で直接還元する製鉄法であることが出土物から判明しています。
粒状水酸化鉄は粘土粒に水酸化鉄が皮膜を作ったものでタイ東北部に無尽蔵にあります。
粒状の原料を炭で直接還元する手法はタタラの砂鉄製鉄と同じ。
水酸化鉄が凝集したもの、これが褐鉄鉱です。
平成3年に長野県飯田市で発見された褐鉄鉱(鬼板)で9回の実証実験がおこなわれ、小刀を作ることに成功しています(伊那史学会)。この実験では小型の竪型炉が使われています。
縄文から弥生へかけての人々の流れに乗るものに稲作があります(≒倭人)。
バンドンブロンの製鉄と同系の製鉄法も稲作の流れに乗って弥生時代初期に到来し、水酸化鉄を探す人々も登場していたのではないか。
弥生中期にはいると大陸の内陸系の人々と文化の流入が多くなり、製鉄も輸入がメインとなって南方系製鉄はすたれていった。
そしてはるか後に原料を砂鉄に置き換えて再登場したのがタタラ製鉄ではないかと考えています(倭鍛冶)。
台湾のアミ族の舟、タタラと呼ばれています。タタラ製鉄の炉の呼称に大船、小舟という呼称があります。
DNAと日本人7 (topへ戻る)
JCウイルスという人間に害を与えず家族間で感染するウイルスがあって、このウイルスの種類を調べることで人間集団の状況を知ることができるのだそうです(東京大学 北村唯一氏の病理学研究の副産物)。
日本では
東北でMY亜型が多い(日本特有タイプ、例外的に中南米にもある)。
西日本でCY亜型が多い(日本特有タイプ。
中部地域で双方同数。
沖縄、九州に少数のSC亜型がある(中国華南に特有タイプ)。
鹿児島、長崎、山梨、富山、山形に少数のB1亜型がある(アジア大陸に広汎に分布)。
青森、秋田、宮城、山形に約1割のEU亜型が含まれる(白人に特有タイプ)。
となったそうです。
世界では
中国西部、モンゴルはB1b亜型、中国東北部はB1a亜型
中国華南地方、台湾はSC亜型
ヨーロッパ、北アフリカ、アラビア半島を含む地域はEU亜型(白人に特有タイプ)
アジア、インド、アラビアからギリシャまでを含む広大な地域はB2亜型
アフリカ大陸、インド、アラビア、ギリシャまでを含む地域はAf2亜型
(ギリシャはウイルス亜型においても東西の十字路なのだそうです)
メキシコ北部、メキシコ中部、ペルーではMY亜型。
下図はそれをイメージ化したものです。
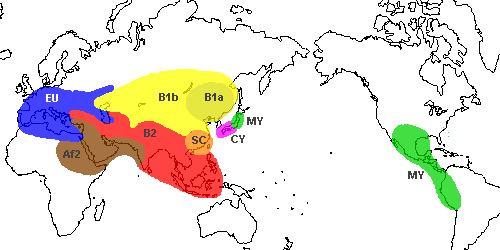
ヨーロッパなどのEU亜型が東北に若干存在すること。
日本にはモンゴルや中国東北部のB亜型がほとんど存在せず、西日本のCY亜型や東日本のMY亜型が日本特有タイプであること。
華南のSC亜型も華南特有タイプで、これは前回書いた9塩基欠失の問題とつながりそうです。
東日本と中央アメリカ大陸原住民が同じMY亜型であることが興味深い。
縄文人が太平洋を渡ったのか??
いまのところシベリア~アラスカ~北アメリカのJCウイルス状況がわからないのでなんともいえません。
(北米経由のユーラシアンが赤道反流に乗ったオーシャノイドと中南米で結合した可能性を考えています)
これらのJCウイルスデータは仮説を補強してくれますが、それが起きた年代がわからないのが残念なところです。
DNAと日本人8 (topへ戻る)
オーストラリア大陸で興味深い骨がでています。
レークマンゴーで発見された3万年前以前とみられる頭骨とカウスワンプで発見されたBC15000頃の頭骨です。
頭骨の発達形状は古い時代のレークマンゴーの方がより進化したタイプで、新しい時代のカウスワンプの方が古く未発達なタイプなのです。
レークマンゴーのDNA分析が行われたそうで、アフリカ起源にはつながらないという結果がでたようですが古い骨のためにDNAが正しく取り出せていないという反論もあり、まだ断定はできないようです。
しかし、オーストラリア大陸には現代人とほぼ同じ人々と、それよりやや古いタイプの2種類の人々が併存していたことに変わりはありません。
これらを単にアフリカ系の人々がやってきた、では説明しにくい。
突然変異をもちだせば簡単ですが、それではなんでもありに近くなってしまいます。
ホモサピエンスとなってアフリカをでてからの14万年前以降が下図です。
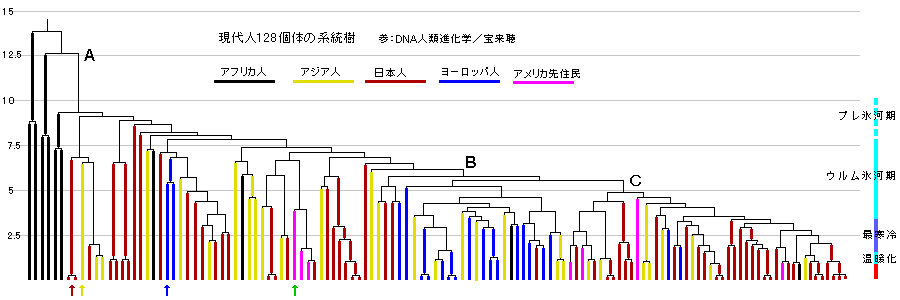
日本人には最古の分岐が含まれ、右側の新しい分岐の人々も多数含まれている。
アジア人もばらついています。各地で複雑な混血(流入混合)があったことを示すのでしょう。
ヨーロッパ人は狭いグループに集中しています。
ヨーロッパ人が近縁の人々だけで構成されていることを示すものと思います。
西アジア、中近東、インド(南部)の人々のデータがほしい、ぜひ分析していただきたいところです。
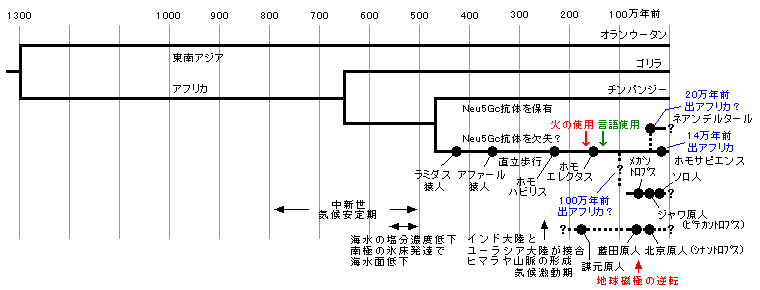
(北京原人やネアンデルタールの登場分岐の年代は明確ではありません)
ゴリラ、人間、チンパンジーのグループは650万年前以降の登場でアフリカ系です。
北京原人やジャワ原人もアフリカ系になっていますが、形態学からの推定だけのようです。
東南アジアのオランウータンが1300万年も前の分岐であることに注目です。
東アジアはアフリカと同等に広大です。原人の進化を消してしまう条件はないとみえます。
北京原人やジャワ原人の系譜が1300万年前に分岐した系譜ではないという証拠はありません。
そこまで遡らずとも100万年前にアフリカを出た原人の子孫が14万年前にアフリカを出た原人の子孫と再会している可能性があります。
モンゴロイドの登場は7万年前以降の単純混血だけではなく、さらに古い人々との結合も含んでいるのではないか。
アフリカ単一発生は同じとしても14万年前よりはるかに古い祖先が混じっている可能性、これを示すのがオーストラリアの骨なのかもしれません。
了
(topへ戻る)
DNAと日本人番外編(血液型)
人類の当初の血液型はみなO型で、これは狩猟民型だそうです。
人口がもっとも多いのもこの型で、肉食型みたい(乳製品は食べません)。
次いで2万5千年ほど前にA型が西アジアあたりに登場して主にヨーロッパ方面へ広がる。
感染症(人口密集地に多い)に抵抗する力が強いのだそうで、農耕民型だそうです。
少し遅れて1万5千年前あたりに中央アジアあたりでB型が登場して東西に広がったのが遊牧民型。
いわゆるモンゴロイドの多くがこのタイプのようです(乳製品をたくさん食べる)。
(ただし、B型を遊牧型と称するのはうまくない可能性があります)
AB型は農耕形と遊牧形の接触による登場とみられるそうで最新版の血液型。
AD500以前のヨーロッパにこの血液型は発見されていません。
A型とB型の得失を両方持ち合わせるために大変複雑な性格になるようです。
AB型の登場が新しいことから、血液型は環境(文化)に千年単位で順応する性質を持つとみえます。
血液型は人種や部族を示すものではなく、文化のありように適応する性質をもつのかもしれません。
(短期間に自然淘汰されるともいえます)
下図は世界の血液型の分布概略です。
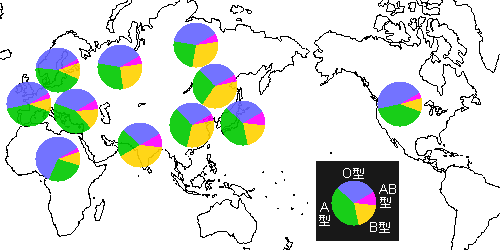
インドでは白人系でもB型が多くヨーロッパでもドイツや東ヨーロッパにはB型が少なからずあるようです。
中国北部や韓国ではB型が多いようですが、日本ではA型が一番多い。
東南アジアは不明ですが、中国南部ではB型が少なくなるようで、環境適応を示すようにみえます。
北米にはB型が少なくヨーロッパに類似なのはヨーロッパからの移住者が多いためでしょう(先住民の血液型を知りたいところです)。
遊牧のB型が多いのはインドと北東アジア。
インドではB形で40%、O形が30%、A形が20%、AB形が10%です。
イラクにはB形が少なくユダヤ人ではB型が多い。
西アジア全般に遊牧のB形が少ないのは早い時期に農耕化したたためでしょうか。
このあたりは一筋縄ではいかない感じ。
日本で一番多いのはA形で40%、O形が30%、B形が20%、AB形が10%です。
日本で農耕のA形が多いのはわかりますが、遊牧のB形というのがどういう経緯で生じたのかむずかしいところ。
縄文の生活様式(木実+魚+雑穀)もB型を生み出すのかもしれません。
わずかですが東日本から東北へゆくほどB形が増え、西へゆくほどA形が増えるようで、これも農耕文化の発達が西からとみれば妥当なところかもしれません。
(topへ戻る)